转载自:小麦研究联盟微信公众号
近日,Food Chemistry在线发表了题为“Selenium and anthocyanins share the same transcription factorsR2R3-MYBandbHLHin wheat”的研究论文。该研究揭示了转录因子R2R3-MYB和bHLH家族在彩色小麦硒与花青素调控网络中的作用。
花青素和硒元素对人和动物均有益,文献报道花青素在蔬菜水果中的含量较高,比如洋葱、大蒜、紫色土豆等(Pyrzynska K., 2009)。结合前人研究结果和课题组对彩色小麦的研究发现,彩色作物多有硒含量较高的特点,但没有直接证据证明富含花青素的彩色作物和硒含量高低是否有直接关联,为此本研究设计了三个实验以揭示硒和花青素之间的关系。
首先是在更广泛的作物类型和品种上进行了硒和花青素的含量测定。本研究选取了6种主要农作物中18个不同颜色品种,测定了可食性部分的硒和花青素含量(图1)。就物种而言,水稻硒和花青素的平均浓度最高(1.41 mg Kg-1, 43.48 mg 100g-1),其次是小麦(0.11 mg Kg-1, 20.51 mg 100g-1)和大麦(0.03 mg Kg-1, 0.43 mg 100g-1)。6种作物的彩色品种的可食性部分均表现出硒与花青素含量高于白皮品种的现象。结合本实验和前人研究结果,彩色品种的硒含量较高可能是一个普遍现象。
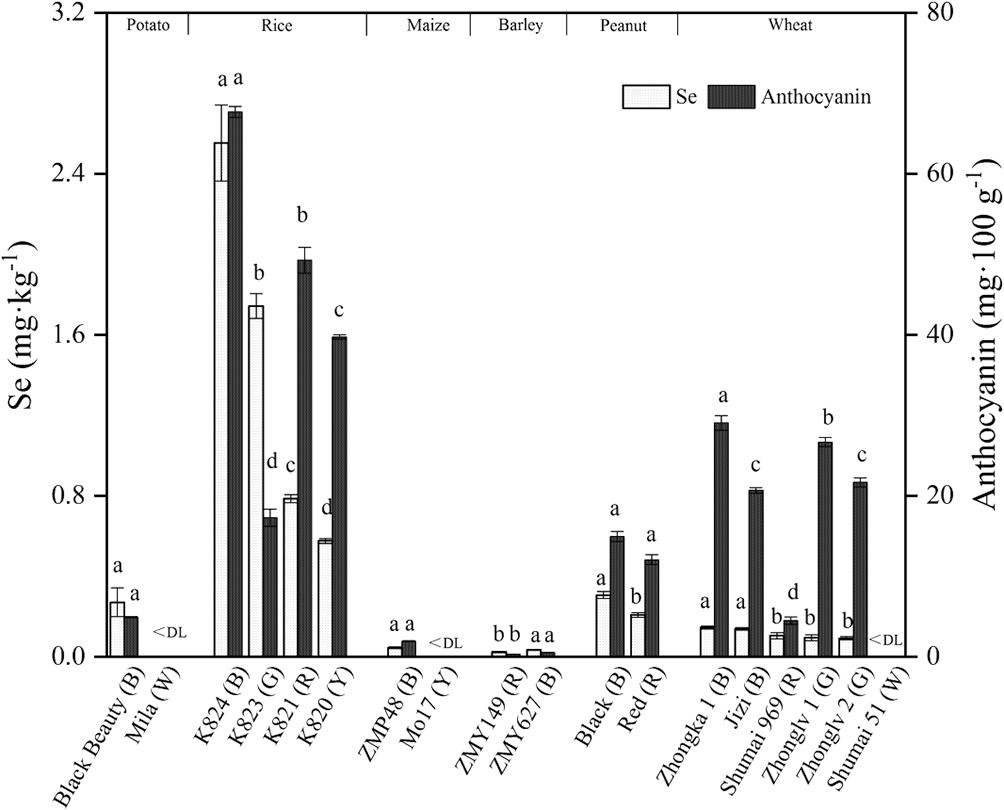
图1 不同作物品种的硒含量及花青素含量
接下来通过对不同颜色的水稻和小麦施加外源硒,分析不同颜色的品种对硒吸收利用效率(图2)。结果表明在不同浓度的硒处理条件下,无论是水稻还是小麦,花青素含量高的品系,其硒吸收效率在同一物种中也较高,颜色最浅白色小麦品种的硒吸收效率最低。不过在同一物种内,硒的吸收效率很大程度上取决于基因型,比如K818和K824均为黑色水稻品种,但二者的硒吸收效率有显著差异。为进一步验证硒与花青素间的关联,我们通过对硒处理后彩色小麦不同种类花青素含量的检测,结果表明,硒不仅对花青素含量产生正向影响,还影响不同类型花青素的积累,其中芍药色素的合成显著受到硒的影响。综上所述,施硒可以同时提高水稻和小麦的花青素和硒含量,其中彩色水稻和小麦有更高的硒吸收效率。
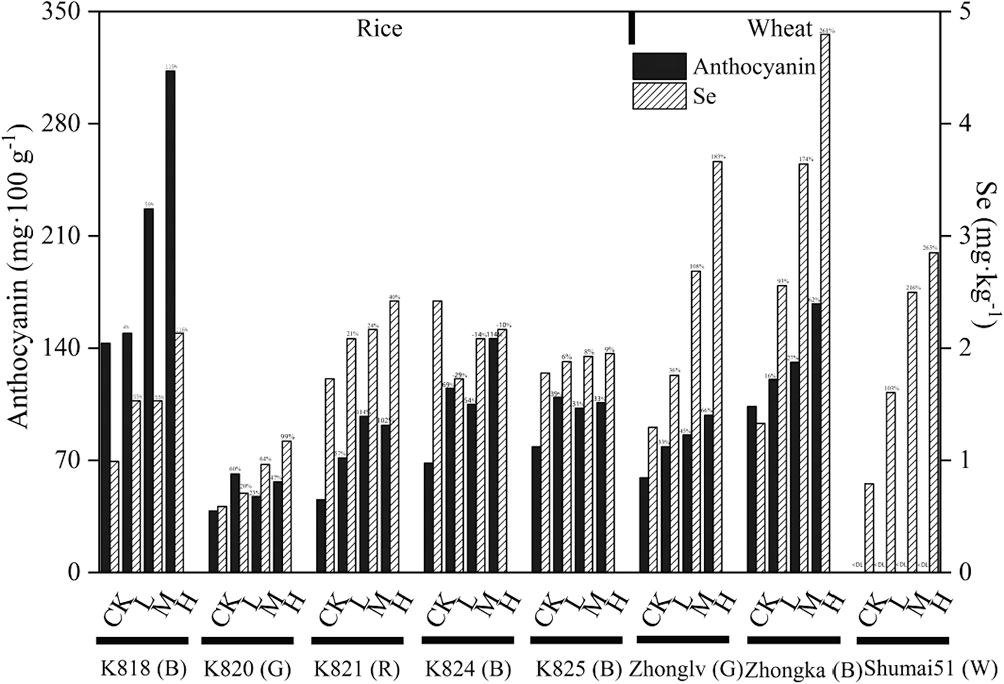
图2 不同硒处理下水稻和小麦中硒和花青素的含量
为进一步明确彩色作物硒吸收效率较高的原因,以彩色小麦和白皮小麦为材料,在外源喷硒条件下,利用转录组测序挖掘调控硒和花青素的关键因子。KEGG富集分析表明,硒诱导上调表达的基因主要集中于苯丙氨酸生物合成、寿命调节和生物信号转导等(图3),其中苯丙氨酸合成途径与花青素代谢途径密切相关,这一结果证明了硒影响了花青素合成途径;上调基因同样富集于与硫代谢途径相关的甘氨酸、丝氨酸和苏氨酸代谢途径以及谷胱甘肽代谢途径。分析不同颜色小麦品种硒处理后相关通路上调基因表达数量,发现转运因子和硫相关基因最多,这也验证了前人关于硒酸盐通过硫代谢转运进入细胞的结论(El Mehdawi A F., et al., 2018 ; Feist, et al., 2001)。综合转录组分析结果表明,即在硒诱导下,同时存在花青素/硫代谢相关的基因富集的现象,解释了彩色作物富硒的可能原因。硒除了通过硫转录因子进行转运,硒也可以通过磷转运因子进入植物细胞。前人研究表明,R2R3-MYB转录因子和bHLH转录因子在磷胁迫反应中均占据重要地位(Gu, M., et al. 2017 ; Yang et al. 2014)。众所周知,R2R3-MYB和bHLH转录因子也参与了花青素的合成途径,从而R2R3-MYB和bHLH在磷代谢途径上将硒与花青素关联起来。随后通过对硒处理后不同颜色的小麦的差异表达基因进行筛选,发现彩色小麦花青素合成的关键转录因子R2R3-MYB和bHLH及在硒诱导下表达量较白色小麦显著提高并通过RT-qPCR得到了验证,表明转录因子R2R3-MYB和bHLH家族或为硒与花青素的调控网络的共同调控因子。综上可得,我们推测出硒与花青素间的可能的调控通路如图4所示。
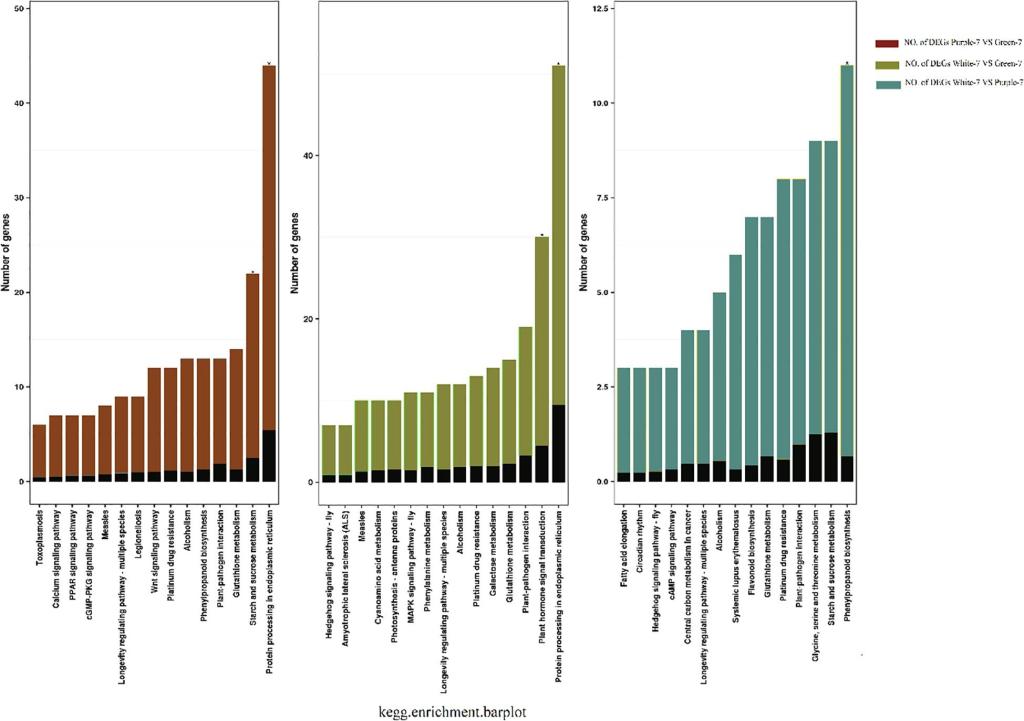
图3 不同粒色小麦品种之间的KEGG通路分析直方图
硫对小麦品质有重要作用,在小麦籽粒中LMW,α+β-麦醇溶蛋白和γ-麦醇溶蛋白是富硫蛋白(Zhao, et al., 1999),本实验也测定了外源硒对蛋白组分的影响,硒主要增加了LMW-GS的含量,不能提高α+β-和γ-麦醇溶蛋白的含量,表明硒与含硫的贮藏蛋白的流向并不完全一致。
本研究揭示了彩色小麦中硒与花青素间的关联,初步探究了硒与花青素互作调控网络中的共同关键因子,为培育富硒富花青素的小麦品种提供了重要的理论依据。下一步我们将继续利用已经纯合的小麦籽粒颜色突变体分析硒和花青素之间的直接关系。
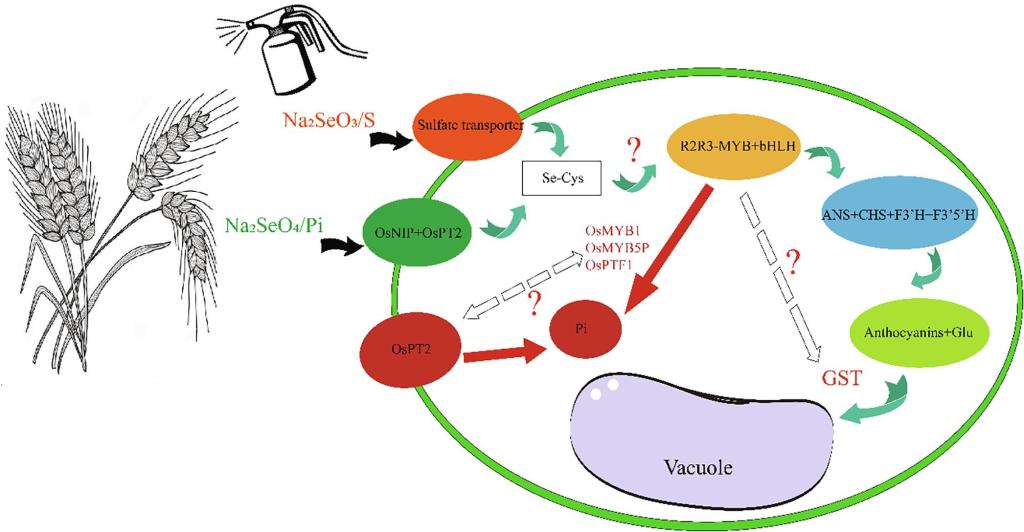
图4 硒与花青素间的调控通路
本研究得到了国家自然科学基金(批准号31971939,31671677),四川省科学技术厅国际科学技术合作与交流计划(2019YFH0063)和四川省重点研究与开发计划(17ZA0315)的支持。
参考文献
[1] Pyrzynska K.. Se speciation in enriched vegetables.Food Chemistry, 2009, 114(4), 1183–1191.
[2] El Mehdawi A F., Jiang Y , Guignardi Z S , et al. Influence of sulfate supply on selenium uptake dynamics and expression of sulfate/selenate transporters in selenium hyperaccumulator and nonhyperaccumulator Brassicaceae[J].New Phytologist, 2018, 217(1): 194-205.
[3] Feist, Laura J., David R., Parker. Ecotypic variation in selenium accumulation among populations of Stanleya pinnata.New Phytologist, 2001, 149(1): 61-69.
[4] Gu, M., Zhang, J., Li, H., Meng, D., Li, R., Dai, X. Maintenance of phosphate homeostasis and root development are coordinately regulated byMYB1, anR2R3-type MYBtranscription factor in rice.Journal of Experimental Botany, 2017, 68(13), 3603–3615.
[5] Yang, W. T., Baek, D., Yun, D.-J., Hwang, W. H., Park, D. S., Nam, M. H., Kim, D. H. Overexpression ofOsMYB4P, anR2R3-type MYBtranscriptional activator, increases phosphate acquisition in rice.Plant Physiology and Biochemistry, 2014, 80, 259–267.
[6] Zhao, F-J., Salmon, S. E., Withers, EJ. A, et al., Variation in the breadmaking quality and rheological properties of wheat in relation to sulfur nutrition under field conditions.Journal of cereal science,1999,30: 19-31